基因表达的调控
遗传信息(基因信息)由DNA转录为RNA、再由RNA翻译为蛋白质,通常称为基因的表达。
基因表达的调控一般表现为正性调控(positive regulation)和负性调控(negative regulation),前者使基因表达量增加,后者则是使基因表达量减少。
一、基因表达的一般特点
同一个体所有细胞都具有相同的基因组,携带个体生存、发育、活动和繁殖所需要的全部遗传信息。但生物基因组的遗传信息(基因)不是同时全部都表达出来,即使是极其简单的生物(如最简单的病毒),其基因组所含的全部基因也不是以同样的强度同时表达。
(一)基因表达具有时间性和空间性
基因表达具有严格的时间和空间特异性,这是由基因的启动子等调控序列与调节蛋白相互作用决定的。
时间特异性(temporal specificity):按照功能需要,某一特定基因表达严格按照一定的时间顺序发生。
阶段特异性(stage specificity):多细胞生物从受精卵到组织、器官形成的各个不同发育阶段,都会有不同的基因严格按照自己特定的时间顺序开启或关闭,表现为分化、发育阶段一致的时间性。(比如兼性异染色质)
空间特异性(spatial specificity):在个体某一发育、生长阶段,同一基因产物在不同的组织器官中表达多少是不一样的。一种基因产物在个体的不同组织或器官中表达,即在个体的不同空间出现。
组织特异性(tissue specificity):不同组织细胞中不仅表达的基因数量不相同,而且基因表达的强度和种类也各不相同。
(二)基因表达有组成性表达和可诱导/阻遏表达两种方式
1.组成性表达
组成性表达(constitutive expression)是指不太受环境变动而变化的一类基因表达。其中某些基因表达产物是细胞或生物体整个生命过程中都持续需要且必不可少的,这类基因称为管家基因(housekeeping gene)。
组成性基因表达也不是一成不变的,其表达强弱是受一定机制调控的。
2.适应性表达
适应性表达(adaptive expression)指环境的变化容易使其表达水平变动的一类基因表达。
应环境条件变化基因表达水平增高的现象称为诱导(induction),这类基因被称为可诱导的基因(inducible gene);相反,随环境条件变化而基因表达水平降低的现象称为阻遏(repression),相应的基因被称为可阻遏的基因(repressible gene)。
改变基因表达的情况以适应环境,在原核生物、单细胞生物中尤其显得突出和重要,因为这些细胞的生存环境经常会有剧烈的变化。
例如,周围有充足的葡萄糖,细菌就可以利用葡萄糖作为能源和碳源,不必更多去合成利用其他糖类的酶类;当外界没有葡萄糖时,细菌就要适应环境中存在的其他糖类(如乳糖、半乳糖、阿拉伯糖等),开放能利用这些糖的酶类基因,以满足生长的需要。
二、原核基因的表达调控
转录调控是原核生物基因表达调控的关键步骤。原核生物大多数基因表达调控是通过操纵子机制实现的。
操纵子(operon)由结构基因和表达调控元件组成,是原核细胞中最常见的表达调控单位。操纵子机制在原核基因表达调控中具有普遍意义。原核基因表达调控最典型的代表是大肠杆菌(E.coil)乳糖操纵子(lac operon)调控模式。在此,以乳糖操纵子为例说明原核基因的表达调控机制。
大肠杆菌乳糖代谢相关基因组成乳糖操纵子。大肠杆菌的乳糖操纵子含LacZ、LacY和LacA三个结构基因,分别编码β-半乳糖苷酶(β-galactosidase)、β-半乳糖苷通透酶(permease)和β-半乳糖苷乙酰转移酶(transacetylase),此外还有一个操纵序列Olac、一个启动序列Plac及一个调节基因LacI(图9-16)。在启动子Plac上游还有一个分解(代谢)物激活蛋白(catabolite activator protein,CAP)的结合位点,与cAMP结合的CAP能结合到CAP结合位点以增强乳糖操纵子的转录活性。
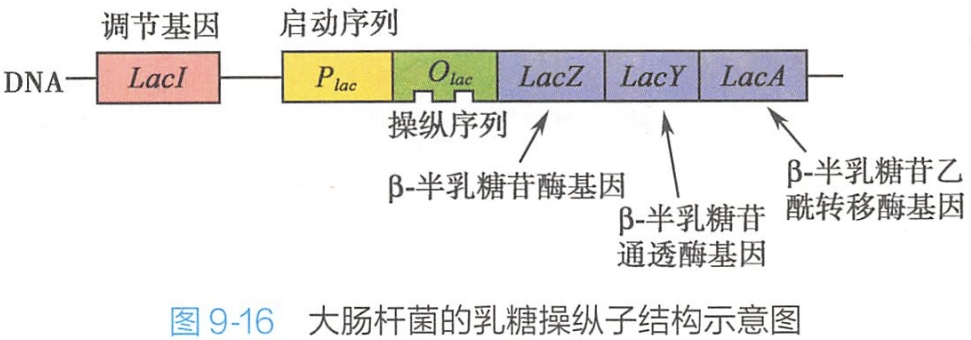
由Plac序列、Olac序列和CAP结合位点共同构成乳糖操纵子的调控区,三个酶的编码基因都由同一调控区调节,实现基因产物的协调表达。
1.环境中没有乳糖
LacI在其自身的启动子控制下,表达产生阻遏蛋白,阻遏蛋白与操纵序列Olac结合,阻碍了RNA聚合酶与启动子Plac的结合,阻止了基因的转录启动。乳糖操纵子处于阻遏状态。
2.有乳糖存在时
乳糖受β-半乳糖苷酶的催化转变为半乳糖,半乳糖与阻遏蛋白结合,使其构象改变,失去与Olac的亲和力,与Olac解离,基因转录开放。
3.有葡萄糖或葡萄糖乳糖共同存在时
细菌首先利用葡萄糖才是最节能的。这时,葡萄糖的存在使细菌内的cAMP浓度降低,阻碍cAMP与CAP结合,从而抑制乳糖操纵子转录,使细菌只能利用葡萄糖。
三、真核基因的表达调控
由于核膜的存在,真核细胞基因的转录与蛋白质的翻译分别在细胞内不同的区域进行。真核细胞基因表达调控可以在转录水平、RNA加工水平、RNA转运水平、RNA降解水平、翻译水平和蛋白质活性水平上进行(图9-17)。
对大多数基因来说,转录水平是最重要的控制点。
此外,在某些细胞中DNA水平的变化如染色质上基因拷贝数的变化、DNA重排、DNA甲基化等也会影响基因的表达,这将在第十五章细胞分化中介绍。(太难了没必要,在分子生物学表观遗传再看吧) #生化
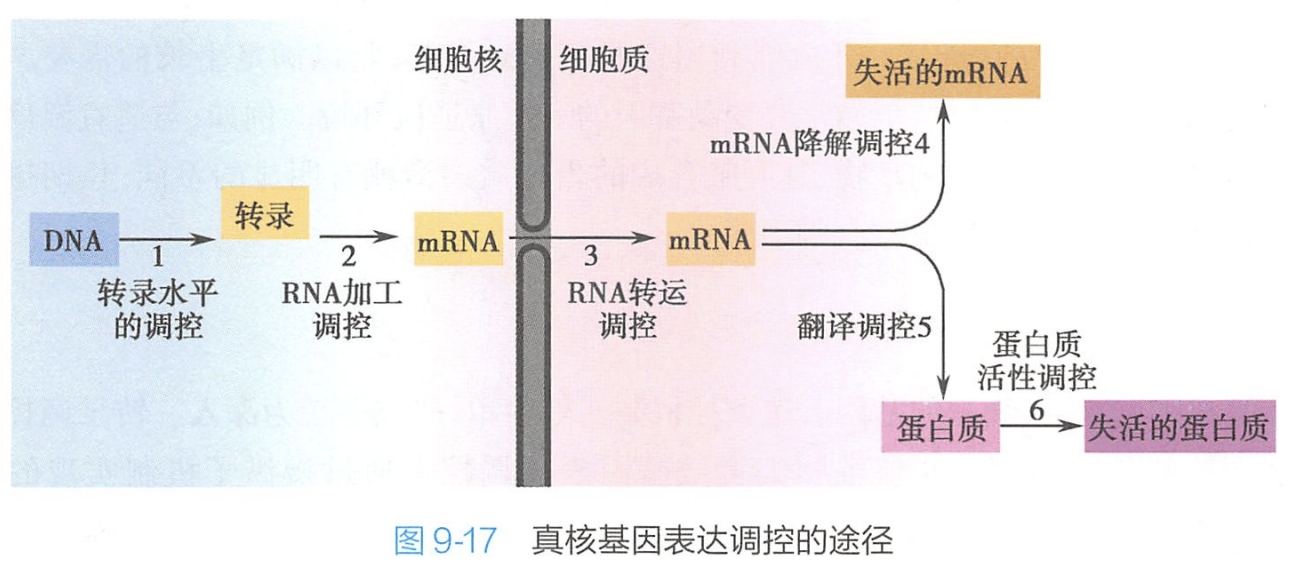
(一)转录水平调控
是真核细胞基因表达的主要控制点。转录水平调控主要是对基因表达转录起始的调节。真核基因具有复杂的调控区,它包括启动子区和其他能调节基因表达的转录因子和基因调节蛋白结合位点。影响基因表达的各种因子通过与 DNA调控区的结合而增强或抑制基因的表达。图9-18显示了真核细胞基因表达调控区的一般结构。
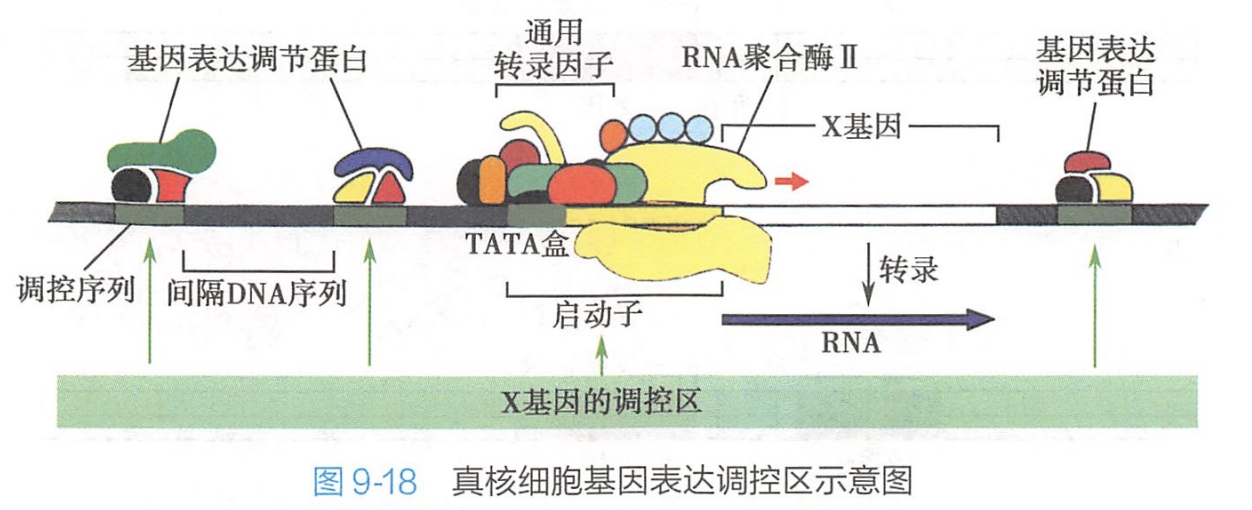
真核细胞中存在多种基因表达调节蛋白,它们能促进或抑制基因的转录,有些基因表达调节蛋白是特异性的DNA结合蛋白,它们能够和靶基因相邻的DNA序列结合;还有一些基因表达调节蛋白虽然不能直接与DNA结合,但它们可通过与其他基因表达调节蛋白的相互作用调控基因转录。
通常将基因表达调节蛋白称做反式作用因子(trans-acting factor),将它们所识别的DNA序列叫做顺式作用元件(cis-acting element)。具体地讲,顺式作用元件是对基因表达有调节活性的DNA序列,其活性只影响与其自身同处在一个DNA分子上的基因,这种DNA序列一般不编码蛋白质,多位于基因旁侧或内含子中。基因表达的调节是通过反式作用因子与顺式作用元件相互作用而实现的。
1.顺式作用元件
根据顺式作用元件在基因组中的位置、转录激活作用的性质及发挥作用的方式,可将真核基因的顺式作用元件分为启动子、增强子(enhancer)和沉默子(silencer)。启动子是决定细胞基因转录起始、能被RNA聚合酶所识别并结合的特异性DNA序列,它是基因准确和有效地进行转录所必需的结构(详见本章第一节)。
增强子是一种能增强真核细胞某些启动子功能的调节序列,不具有启动子的功能,但能增强或提高启动子的活性。
- 增强子在DNA双链中没有5'与3'固定的方向性,作用不受序列方向性的限制。增强子在所调控基因的上游或下游均可发挥作用,但大多位于上游;下游内含子中,乃至下游最后一个外显子以外的序列也可含有增强子。
- 增强子在距离启动子相对较远(距启动子数kb至数十kb)时也能发挥作用。
- 增强子一般有组织或细胞特异性,
- 但对启动子的影响无严格的专一性。基因重组实验证明,同一增强子可影响不同类型的启动子,真核生物增强子也可影响原核生物的启动子(图9-19)。
在顺式作用元件中还存在一种与增强子作用相反的沉默子。沉默子是基因表达的负性调控元件。当特异蛋白因子与其结合时,对基因转录起阻遏作用。沉默子的作用可不受序列方向的影响,能远距离发挥作用,并可对异源基因的表达起作用。
2.反式作用因子
真核基因表达的调节蛋白又称转录调节因子或转录因子。绝大多数真核转录调节因子由它的编码基因表达后,通过与特异的顺式作用元件的识别、结合(即DNA-蛋白质相互作用)反式激活另一基因的表达。
RNA聚合酶和转录因子就是反式作用因子。能直接结合DNA序列的反式作用因子是少数,但不同的反式作用因子之间可以相互作用,因而目前认为多数转录因子是通过蛋白质-蛋白质间作用与DNA序列联系并影响转录效率的,转录因子之间或转录因子与DNA的结合都会引起DNA构象的变化,从而调节转录。
作为蛋白质的转录因子从功能上分析其结构,可包含有不同区域:
- DNA结合域(DNA binding domain):多由60~l00个氨基酸残基构成的几个亚区组成。不与DNA直接结合的转录因子没有DNA结合域,但能通过转录激活域直接或间接作用于转录复合体而影响转录效率。与DNA结合的转录因子大多以二聚体形式起作用,其DNA结合域常见有以下几种:
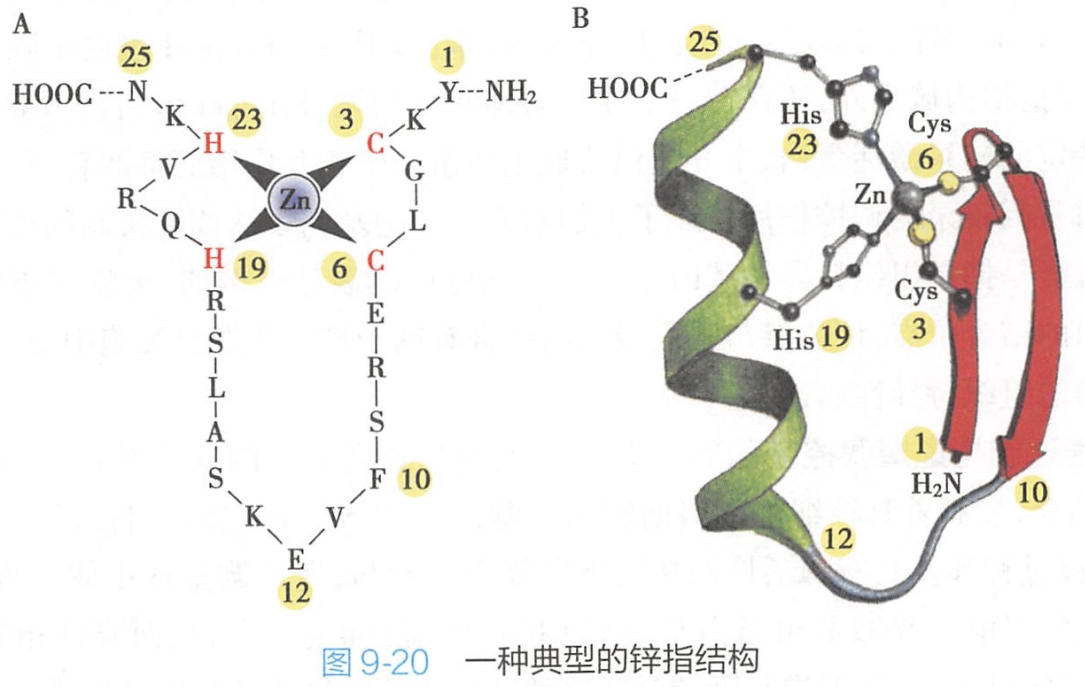
- 锌指结构(zinc finger):是最常见的DNA结合域,它由约23个氨基酸残基组成,其中的半胱氨酸和组氨酸通过配位键与锌原子结合,这样形成一个以锌原子为中心的“指”状结构,称为锌指结构。锌指结构的N-末端形成β-片层,C-末端形成α-螺旋,α-螺旋能够与DNA大沟相结合(图9-20)。一个转录因子中常常含有多个串联重复的锌指结构,如与GC盒结合的转录因子SP1中就含有3个重复的锌指结构,这些锌指结构中的3个α-螺旋恰好等于DNA大沟的一圈。
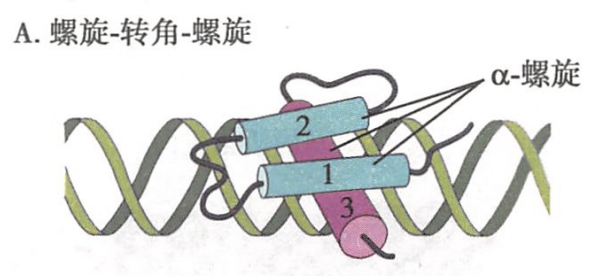
- 螺旋-转角-螺旋(helix--turm-helix,HTH):如图9-21A所示,这类结构域通常由3个a-螺旋结构组成,其中一个螺旋与DNA大沟结合,另两个螺旋则位于其上面起稳固作用。例如,参与大肠杆菌乳糖操纵子调控的CAP蛋白以及真核细胞中的同源异形结构域(homeodomain)蛋白中就含有HTH结构域。
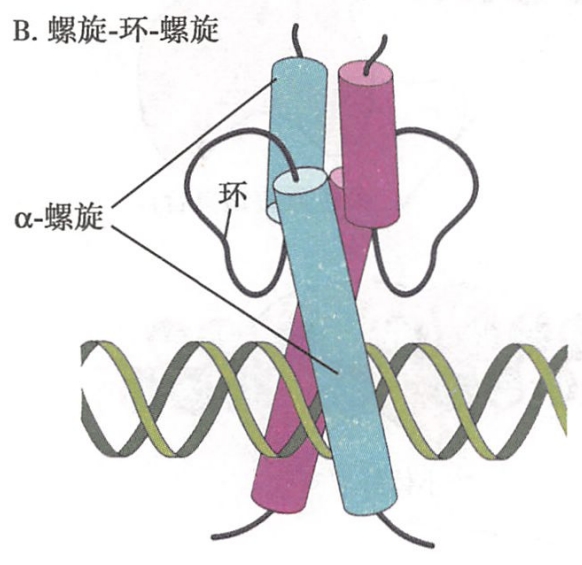
- 螺旋-环-螺旋(helix-loop-helix,HLH):这类结构至少有两个α-螺旋,螺旋间由短肽段形成的环连接,两个具有HLH结构的转录因子以二聚体形式相连,两个较长的α-螺旋刚好能够嵌入DNA的大沟(图9-21B)。例如,参与肌细胞生成的转录因子MyoD蛋白就含有HLH结构域。
- 亮氨酸拉链(leucine zipper):该结构的特点是蛋白质分子的肽链上每隔6个氨基酸就有一个亮氨酸残基,结果导致这些亮氨酸残基都在α-螺旋的同一个方向出现,两个这样的α-螺旋肽链单体就能通过亮氨酸残基以疏水键结合形成二聚体。该二聚体另一端的肽段形成“Y”形的结构(富含碱性氨基酸残基)与DNA大沟结合(图9-21C)。
- 锌指结构(zinc finger):是最常见的DNA结合域,它由约23个氨基酸残基组成,其中的半胱氨酸和组氨酸通过配位键与锌原子结合,这样形成一个以锌原子为中心的“指”状结构,称为锌指结构。锌指结构的N-末端形成β-片层,C-末端形成α-螺旋,α-螺旋能够与DNA大沟相结合(图9-20)。一个转录因子中常常含有多个串联重复的锌指结构,如与GC盒结合的转录因子SP1中就含有3个重复的锌指结构,这些锌指结构中的3个α-螺旋恰好等于DNA大沟的一圈。
- 转录激活域(activating domain):常由30~l00个氨基酸残基组成,该结构域包含富含酸性氨基酸、富含谷氨酰胺、富含脯氨酸等不同种类。
- 连接区:即连接上两个结构域的部分。
总之,反式作用因子能够通过识别启动子、启动子附近和增强子等顺式作用元件中的特异靶序列并与之结合(即蛋白质-DNA相互作用),以及通过蛋白质-蛋白质的相互作用,最终影响RNA聚合酶活性,从而对基因表达发挥正调控或负调控作用。通常把发挥正调控作用的反式作用因子称为转录激活蛋白(activator)。
3.阻遏蛋白
真核细胞中除存在发挥正性调控作用的转录激活蛋白之外,还有一类阻遏蛋白(repressor,也称抑制子)也参与了真核细胞基因表达的负性调控。
与原核细胞中起同样作用的分子一样,真核阻遏蛋白通过与特定的DNA序列结合而抑制转录,即都有DNA结合域。
- 一类阻遏蛋白:通过阻止激活蛋白与启动子或增强子的结合,抑制转录。
- 启动子:其中,某些阻遏蛋白通过干扰其他转录因子与DNA的结合来发挥抑制作用,例如,在转录起始位点附近结合的阻遏蛋白可以阻止RNA聚合酶或通用转录因子与启动子的相互作用;
- 增强子:另一些阻遏蛋白则通过与转录激活蛋白竞争结合DNA分子中的调控序列发挥作用。这些阻遏蛋白含有和激活蛋白相同的DNA结合域,但是阻遏蛋白没有转录激活域,无法激活转录。
- 另一类阻遏蛋白,称为活性阻遏蛋白(active repressor)。与那些仅仅干扰转录激活蛋白结合DNA的阻遏蛋白不同,它们虽然也有DNA结合域,但主要靠功能结构域通过蛋白质-蛋白质的相互作用来抑制转录。例如果蝇的Krüppel基因,其表达的蛋白质即为活性阻遏蛋白。Krüppel蛋白中除了含有一个能够与DNA结合的锌指结构域之外,还存在一个独立的抑制结构域,Krüppel蛋白与DNA结合后,依赖其抑制结构域与特定的转录激活蛋白、中介蛋白或通用转录因子相互作用而抑制转录。
阻遏蛋白一个重要的功能可能是抑制组织特异性基因在不适当细胞中的表达。例如,免疫球蛋白基因的增强子中含有阻遏蛋白的结合位点,该位点与阻遏蛋白的结合有助于抑制非淋巴细胞中免疫球蛋白的表达,确保免疫球蛋白的组织特异性表达。
4.染色质重塑
染色质是高度有序的紧密结构,限制了转录因子对DNA的接近和结合,控制着真核细胞基因的转录。基因表达激活首先需要将致密压缩的染色质/核小体舒展开来,该过程涉及具有酶活性的功能蛋白复合体参与,通过调整核小体结构,中和组蛋白碱性氨基酸残基上的正电荷来减弱组蛋白与DNA(携负电荷)间的结合,从而降低相邻核小体间的聚集,增加转录因子的进入,最终促进基因转录。这种染色质结构的动态变化过程通常称为染色质重塑(chromatin remolding)。
染色质重塑是基因表达调控的主要方式之一。目前认为引起染色质重塑的方式主要有以下两种(图9-22):
(1)依赖ATP的物理性修饰
即以ATP水解释放的能量,使组蛋白和DNA的构象发生局部改变。该过程主要通过依赖ATP的染色质重塑复合体或染色质重建子(remodeler)来完成,这些复合体是一种以ATP酶为催化中心的多种蛋白亚基复合体,如SWI/SNF复合体,能够被DNA结合的激动子(activator,也称基因活化蛋白)或抑制子(repressor)招募至启动子部位(应该是转录激活蛋白和阻遏蛋白的意思吧),借ATP水解的能量移动核小体的位置使DNA序列暴露或被掩盖,而参与基因表达调控。
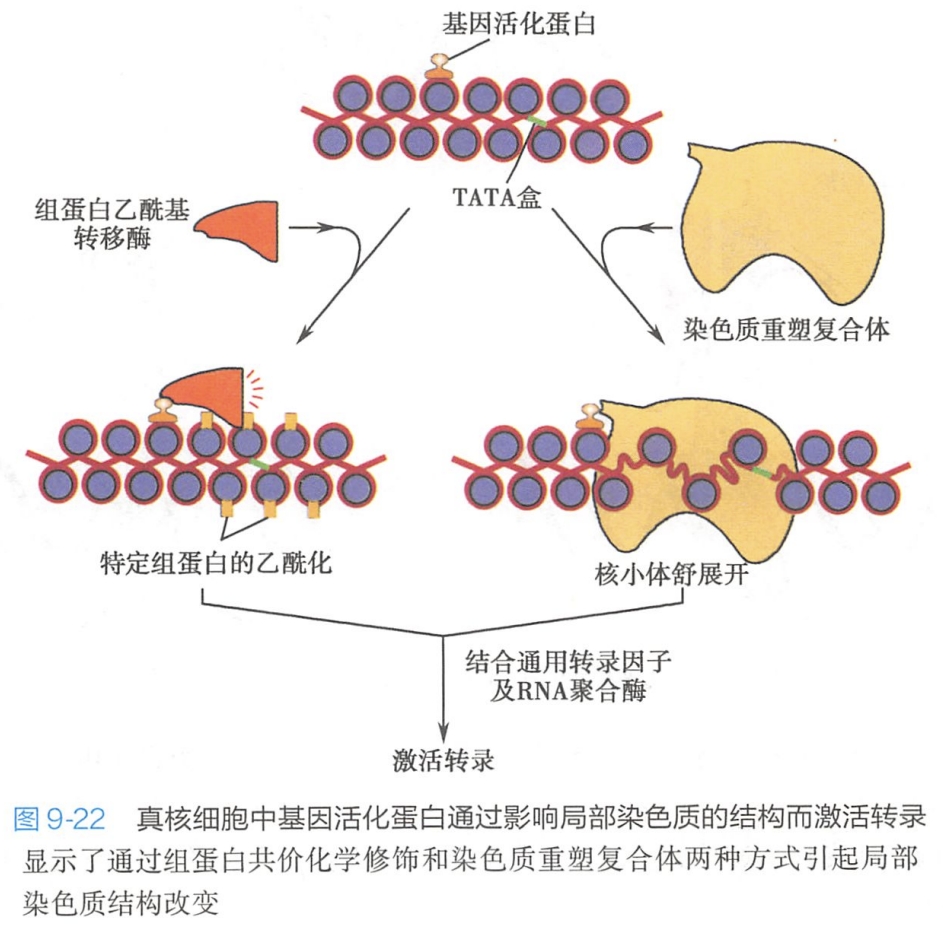
(2)组蛋白共价化学修饰
组蛋白的结构是动态变化的,这种变化影响了染色质结构的构型,从而调节基因的表达。组蛋白修饰导致基因转录或沉默的机制与其引起的染色质重塑密切相关。组蛋白N-端尾部的氨基酸修饰直接影响了核小体的结构,进而影响到转录起始复合体是否易于同启动子部位的DNA结合。
组蛋白结构的改变源于组蛋白中被修饰的氨基酸。核小体的核心组蛋白(H2A、H2B、H3和H4)是一类小分子量的强碱性蛋白,它们均由球状结构域(外周被146bp大小DNA包绕)和从核小体表面伸出的位于蛋白N-端的“组蛋白尾巴”组成。组蛋白特别是H3和H4尾部的氨基酸残基能够被化学修饰,包括组蛋白的乙酰化(acetylation)、甲基化(methylation)、磷酸化(phosphorylation)、泛素化(ubiquitination)、sumo化(sumoylation)、多聚ADP-核糖基化[poly(ADP-ribosyl)ation]等。
组蛋白中被修饰氨基酸的种类、位置和修饰类型以及各种修饰在时间、空间上的组合与生物学功能的关系被称为组蛋白密码(histone code),它决定了染色质转录活跃或沉默的状态。图9-23描述了组蛋白修饰的一般概念和常见标记。

以组蛋白乙酰化为例,说明组蛋白修饰影响基因转录的机制。组蛋白乙酰化多发生于H3和H4氨基酸的赖氨酸残基。在组蛋白乙酰基转移酶(histone acetyltransferase,HAT,也称乙酰化酶)作用下,于组蛋白N-端尾部的赖氨酸加上乙酰基,称为组蛋白乙酰化。组蛋白N-端的赖氨酸残基乙酰化会移去正电荷,降低组蛋白和DNA之间的亲和力,使得RNA聚合酶和通用转录因子容易进入启动子区域(图9-22)。因此,在大多数情况下,组蛋白乙酰化有利于基因转录。低乙酰化的组蛋白通常位于非转录活性的常染色质区域或异染色质区域。
其具体机制是:基因激活因子结合于特定上游激活序列(UAS)并招募组蛋白乙酰化酶,催化附近的组蛋白乙酰化,促进基因激活;而结合于上游抑制序列(URS)的转录抑制因子则招募组蛋白去乙酰化酶(histone deacetylase,HDAC),催化附近的组蛋白去乙酰化,抑制转录。
5.DNA甲基化
真核DNA分子中的胞嘧啶碱基约有5%被甲基化修饰为5-甲基胞嘧啶。胞嘧啶碱基的甲基化修饰常发生在基因上游调控序列的CpG(岛)富含区。(p是中间磷酸,应该与GC盒不同吧)
处于活性染色质区、呈现转录活性状态的基因的CpG序列一般总是低甲基化的;而不表达或处于低表达水平的基因的CpG序列则总是高甲基化的。
实际上,DNA胞嘧啶的甲基化与去甲基化修饰是一个动态的过程,在真核基因的表达调控中发挥着重要作用。DNA甲基转移酶(DNA methyltransferase, DNMT)催化甲基与胞嘧啶的共价结合,而去甲基化酶(demethylase)则负责去除5-甲基胞嘧啶上的甲基。
6.ncRNA
除了转录调节蛋白以外,一些非编码RNA分子,如miRNA等小RNA和lncRNA能调控基因的转录。相对于长链非编码RNA,人们对小RNA(miRNA)的作用机制有较多的认识。在第二章中已经介绍过miRNA在发挥RNA干扰(降解mRNA)中的作用机制(图2-12),在此简述miRNA在调节基因转录中的作用。
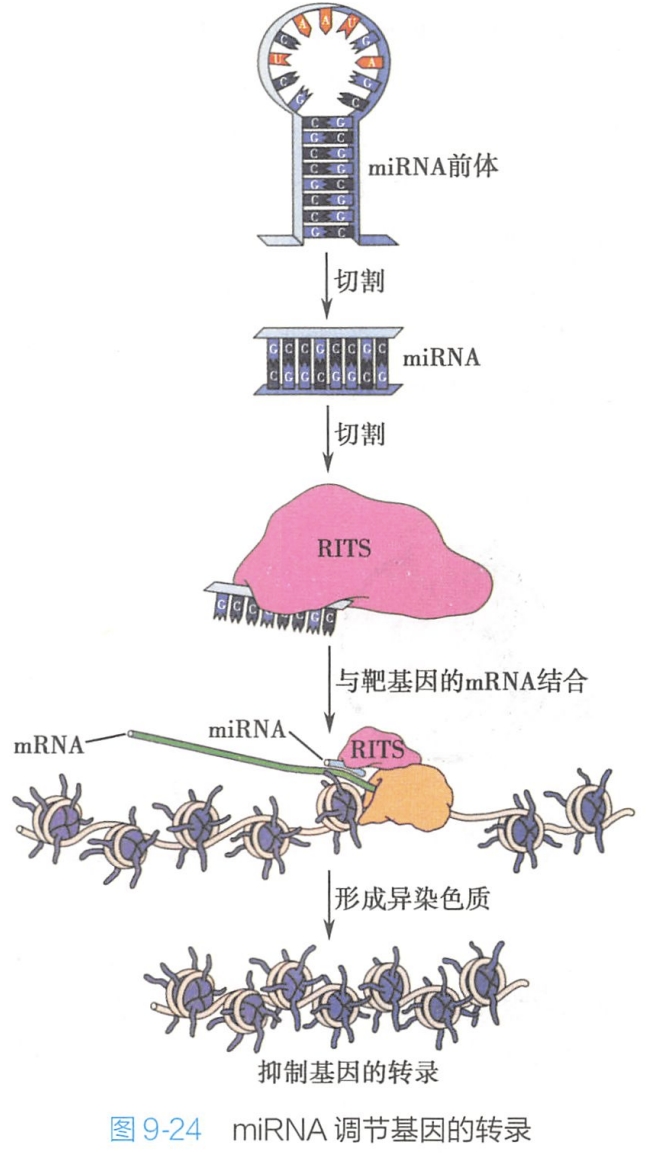
在miRNA发挥转录抑制作用时,miRNA与另一种蛋白质复合体——RITS(RNA-induced transcriptional silencing)复合体结合,解离后的一条miRNA链将RITS复合体引导至同源基因处(很可能是通过碱基配对结合于同RNA聚合酶Ⅱ结合的mRNA上),然后,RITS复合体通过募集组蛋白甲基转移酶,使组蛋白H3的赖氨酸-9发生甲基化,导致异染色质形成,最终抑制基因的转录(图9-24)。
(二)hnRNA水平调控
1.常规剪接
多数真核细胞的mRNA前体分子经剪接去除内含子后仅形成一种成熟的mRNA,并经过翻译生成一条相应的多肽链,这种RNA剪接方式称为常规剪接(constitutive splicing)。
2.可变剪接
但许多高等真核细胞的mRNA前体不止含有一个内含子,在特定条件下,剪接发生在两个内含子之间,即某个内含子的 5'端与另一个内含子的3'端进行剪接时,就会删除这两个内含子及其中间的全部外显子或内含子,这称为RNA可变剪接(alternative splicing)(详见第十五章细胞分化,图15-14)。
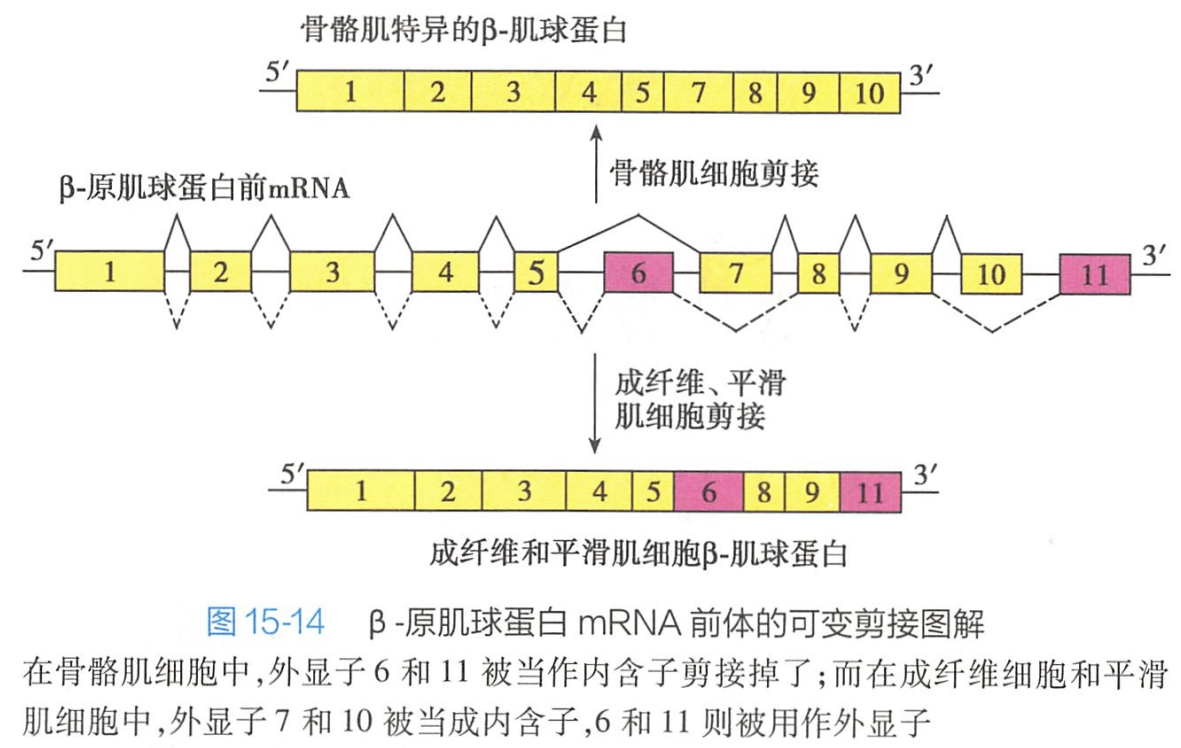
由此,一个基因的mRNA前体经过可变剪接可以产生多种不同的蛋白质,或形成一组相似的蛋白质家族。
3.剪接体和剪接位点
mRNA前体的剪接由被称为剪接体的核酸-蛋白质复合物完成,该复合物主要包括snRNA及富含丝氨酸和精氨酸的相关蛋白。
剪接首先是剪接体识别RNA前体剪接位点,剪接位点的选择受许多顺式作用元件和反式作用因子的调控。反式作用因子可以通过识别正性或负性的顺式作用元件对不同的剪接位点进行选择。
此外,剪接体中的剪接因子也参与了对选择性剪接的调控。
(三)翻译水平调控
与起始因子磷酸化密切相关。真核细胞翻译水平调节点主要在翻译起始阶段和延长阶段,尤其是起始阶段。如起始因子活性的调节、Met-tRNAMet与核糖体小亚基结合的调节、mRNA与小亚基结合的调节等。其中通过磷酸化作用改变起始因子活性这一点备受关注。
蛋白质合成速率的快速变化很大程度上取决于起始水平,通过磷酸化调节起始因子活性对起始阶段有重要的控制作用。如eIF-2α亚单位的磷酸化可以阻碍eIF的正常运行,从而抑制蛋白质合成的起始。eIF-2α亚单位的磷酸化由特异蛋白激酶催化。在病毒感染的细胞中,细胞抗病毒机制之一即是通过双链RNA(double-stranded RNA,dsRNA)激活一种蛋白激酶,使eIF-2α磷酸化,从而抑制蛋白质合成的起始。
此外,mRNA的5'UTR(非翻译区)与3'UTR在翻译过程中能够相互依赖、协同作用提高翻译效率。一些蛋白质能够识别RNA的3'UTR,通过干扰3'poly-A尾与5'端帽的联络而减少翻译的起动。